


This article requires a subscription to view the full text. If you have a subscription you may use the login form below to view the article. Access to this article can also be purchased.
- Address for Correspondence: Nadine M. Lerret
, Rush University, nadine_lerret{at}rush.edu
ABSTRACT
The major driving force in the pathogenesis of prediabetes and type 2 diabetes is thought to be chronic and low-grade systemic inflammation. Immune cells are key players in the induction of this chronic inflammation; however, the type of immune cells and mechanisms underlying the rapidly progressing pathogenesis of this disease are unclear. Therefore, we examined whether hyperglycemia alters CD4 T-cell activation, differentiation, and survival. Dendritic cells and T cells were isolated from human peripheral blood and cultured with varying amounts of glucose. Using flow cytometry, we found dendritic cells primed in hyperglycemic conditions induced CD4 T cells to exhibit a more activated phenotype capable of mobilization. This was evident by upregulation of the adhesion molecule CD11a. Additionally, these activated CD4 T cells had a lower degree of proliferation and decreased expression of the apoptotic protein caspase-3 compared with CD4 T cells primed by dendritic cells stimulated with a physiological concentration of glucose. We conclude that hyperglycemia drives CD4 T cells toward an activated immunophenotype, which could be assessed via flow cytometry in the clinic to further stratify patients with prediabetes and type 2 diabetes and potentially predict those patients at highest risk for progression of the disease.
- CFSE - carboxyfluorescein succinimidyl ester
- DC - dendritic cell - HgbA1c - hemoglobin A1c
- IL - interleukin
- PBMC - peripheral blood mononuclear cell
- PBS - phosphate-buffered saline
- diabetes mellitus
- type 2
- CD4-positive T lymphocytes
- inflammation
- hyperglycemia
- prediabetic state
- insulin resistance
INTRODUCTION
Type 2 diabetes is one of the most prevalent chronic metabolic illnesses in the United States, affecting more than 29.1 million individuals to date.1,2 Clinical diagnosis focuses on hyperglycemia, which is characterized by a hemoglobin A1c (HgbA1c) value ≥6.5% and fasting plasma glucose >125 mg/dl.3 In addition, there are at least 86 million people categorized as prediabetic who have elevated HgbA1c values between 5.7%–6.4% and fasting plasma glucose values between 100–125 mg/dl.1⇓-3 Unfortunately, a majority of prediabetic patients progress to type 2 diabetes and have a significant risk of developing complications from this disease, such as diabetic retinopathy, kidney disease, stroke, and cardiovascular disease.2,4 For this reason, early detection of patients with prediabetes or early onset type 2 diabetes is crucial to prevent diabetic-associated complications and comorbidities.
Recently, it has been proposed that hyperglycemia and chronic inflammation play a role in the pathogenesis from prediabetes to type 2 diabetes.4 In support of this, it was found that a majority of patients with type 2 diabetes are overweight or obese, exhibit obesity-linked insulin resistance, and have elevated inflammatory circulating factors.5 Emerging data also suggest a pathological role for CD4 T cells in obesity, insulin resistance, and disease progression to type 2 diabetes. One study found activated CD4 T cells and increased inflammatory cytokines in the kidneys of patients with type 2 diabetes.6 Subsequent clinical research has shown an increase in CD4 T cells from both peripheral blood and adipose tissue of people with type 2 diabetes.7 Recently, it was demonstrated that dendritic cells (DCs) exhibit a more mature phenotype after being cultured in media containing high concentrations of glucose.8 This study suggests that circulating high glucose drives immature DCs toward maturation; however, it did not reveal whether hyperglycemia influences CD4 T-cell priming by DCs and what the downstream effects are on CD4 T-cell phenotype and function when DCs are initially primed in a hyperglycemic environment.
We hypothesized the activated state of hyperglycemic-induced DCs would subsequently alter CD4 T-cell priming. Data herein show the downstream effect of DCs cultured in hyperglycemic conditions manifests as an increase in CD4 T-cell activation. This event is characterized by increased expression of CD11a on CD4 T cells. Additionally, although proliferation of CD4 T cells from hyperglycemic culture conditions was decreased compared with physiological and prediabetic culture conditions, the CD4 T cells from diabetic culture conditions were less likely to undergo apoptosis. Taken together our data provide evidence that hyperglycemia alters the immune phenotype and cell survival of human T cells.
MATERIALS AND METHODS
Cell Separation and Isolation of Peripheral Blood Mononuclear Cells
Isolation of human peripheral blood mononuclear cells (PBMCs) was carried out using Histopaque® (MP Bio) density-gradient centrifugation. Human whole blood used for PBMC isolation was provided as a unit of deidentified whole blood from LifeSource (Chicago, IL). The blood was diluted 1:2 with phosphate-buffered saline (PBS) (MP Bio) and then layered over Histopaque® (MP Bio) and centrifuged at room temperature (20–25 oC) for 30 minutes at 2000 rpm. The interface (buffy coat) was recovered and washed three times with PBS (MP Bio). This study was exempt from institutional review board approval as all blood used was completely deidentified and donors did give their informed consent to donate peripheral blood.
Immature Dendritic-cell and T-cell Isolation From PBMCs
CD14+ and CD3+ cells were isolated from the PBMCs using CD14- and CD3-labeled magnetic beads (Miltenyi Biotec). Separate PBMC fractions were incubated with either CD14 or CD3 beads for 20 minutes at 4 oC and then applied to an MACS magnetic column (Miltenyi Biotec). After washing the column three times with MACS buffer (Miltenyi Biotec), CD14+ immature DCs were eluted (resulting purity >85% as determined by flow cytometry) and CD3+ T cells were eluted (resulting purity 87.6% as determined by flow cytometry) from the column.
Generation of Monocyte-derived DCs
Isolated immature CD14+ DCs (1 × 105/ml) were cultured for 6 days in 6-well tissue culture plates in glucose-free HyCloneTM RPMI media supplemented with 10% fetal bovine serum (Gibco), granulocyte-macrophage colony-stimulating factor (100 ng/ml), interleukin (IL)-4 (50 ng/ml), 1 mM sodium pyruvate (Gibco), 0.1 mM nonessential amino acids (Gibco), and 1% penicillin/streptomycin (Gibco). Cells were incubated at 37 oC with 5% CO2 with media changed every 2 days. On day 6 of culture, DCs were administered varying concentrations of glucose (Fisher Scientific) and incubated for an additional 24 hours to allow sufficient time for glucose stimulation. The conditions included complete absence of glucose or the addition of 5.5 mmol/L (physiologic), 15 mmol/L (prediabetic), or 30 mmol/L (hyperglycemic) glucose concentrations.
DC: T-cell Coculture
Following incubation with or without glucose for 24 hours, naïve autologous CD3 T cells (1 × 106/ml) were added to the cultured DCs. The culture medium was supplemented with IL-2 (80 U/ml; Fisher Scientific). It is possible the DCs utilized all of the glucose before the T cells were exposed; therefore, T cells cocultured with DCs in the absence of glucose provided a valuable internal control. In order to measure proliferation, some experiments utilized CD3 T cells prelabeled with 0.5 mM carboxyfluorescein succinimidyl ester (CFSE; Tonbo Biosciences). CFSE is a fluorescent cell-staining dye used to monitor distinct generations of proliferating cells by dye dilution. Therefore, the more times a cell proliferates, the weaker the fluorescent signal. T cells stimulated with CD3/CD28 beads (Tonbo Biosciences) were plated as a positive proliferation control. CD3/CD28 beads were used to mimic in vivo antigen presentation through T-cell coreceptor (CD3) and costimulatory molecule (CD28) engagement. T cells cultured alone, without DCs or CD3/CD28 beads, were used as a negative proliferation control. The DCs and T cells were cocultured for 7 days before harvesting for flow cytometry assays.
Flow Cytometry Assays
Lymphocytes were stained with fluorochrome-conjugated monoclonal antibodies directed against human CD3 (SK7), CD4 (SK3), CD11b (ICRF44), CD14 (61D3), CD83, and CD45 (HI30), all from Tonbo Biosciences, and CD11a (HI111), and caspase 3 (C92-605) from BD Pharmingen. Flow cytometry was performed on a BD LSRII instrument, and data were analyzed with FlowJo software.
Statistical Analysis
Data were expressed as the mean ± SEM of independent experiments. Statistical significance was determined by Student two-tailed paired t test, assuming equal variances, or ANOVA. P values <0.05 were considered statistically significant.
RESULTS
CD4 T Cells Show Decreased Proliferation When Primed With DCs Exposed to Hyperglycemic Conditions
High glucose was previously shown to induce maturation of human monocyte–derived DCs in culture.8 This was demonstrated by an increase in the costimulatory molecules CD83 and CD86 expression by DCs exposed to 15 mmol/L or 30 mmol/L of glucose compared with DCs exposed to 5.5 mmol/L glucose.8 However, the resulting effect of this induced maturation on T cells has not been explored. To accomplish this, naïve autologous CD3 T cells were cocultured with DCs previously stimulated by physiologic (5.5 mmol/L), prediabetic (15 mmol/L), or hyperglycemic (30 mmol/L) concentrations of glucose. CD3 T cells cocultured with DCs matured in the absence of glucose were included for comparison.
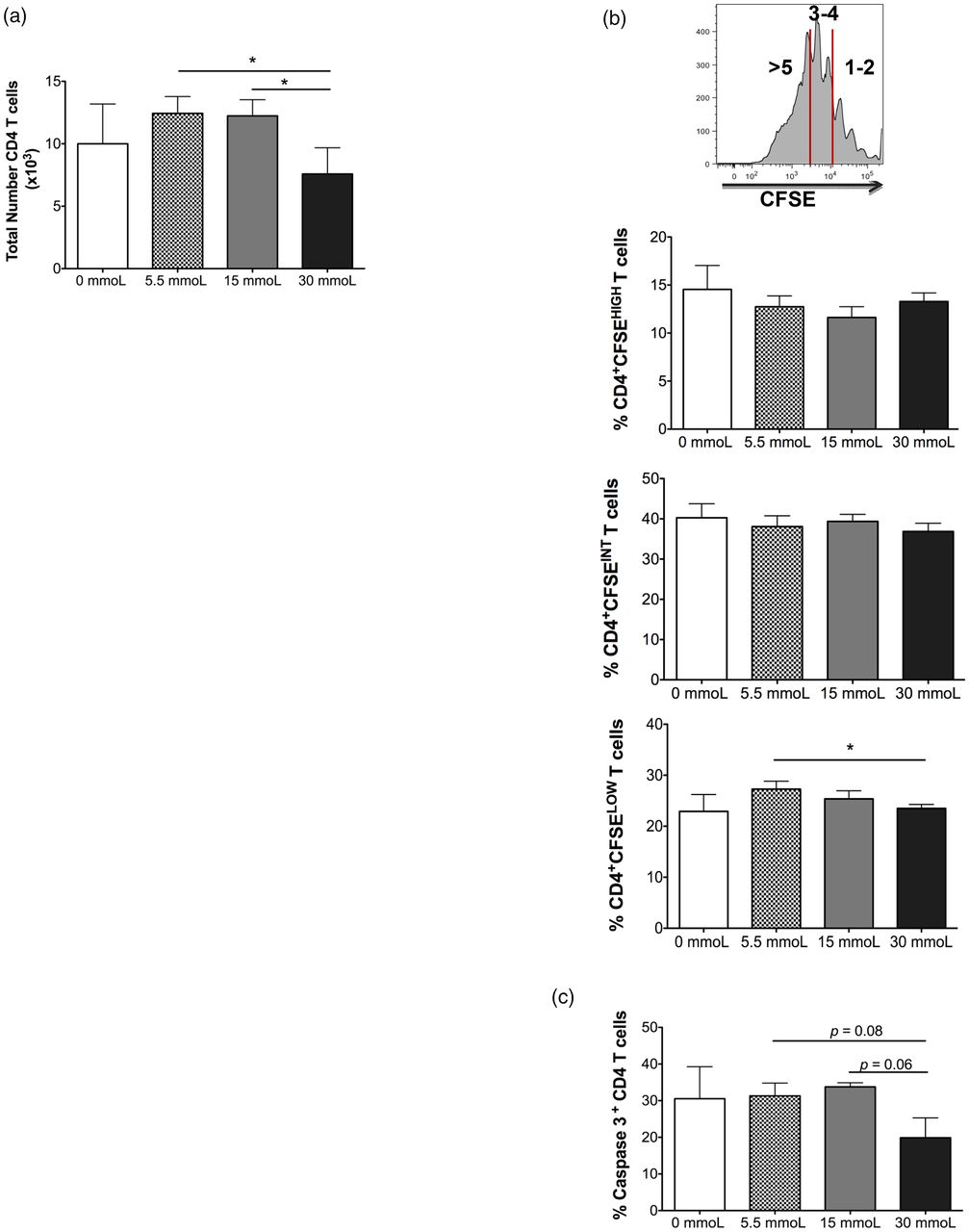
We first analyzed whether there was a difference in the total number of CD3+ CD4 T cells recovered after coculture with DCs matured in different concentrations of glucose. CD4 T cells were specifically chosen as the CD3+ subset to analyze in this study (ie, gated on via flow cytometry) because recent reports demonstrate a pathological role for CD3+ CD4 T cells in obesity and insulin resistance.7,9,10 Seven days post-coculture, the number of CD4 T cells recovered from wells with DCs exposed to the highest concentration of glucose was significantly reduced compared with wells that had DCs matured in physiological glucose (mean, 7.6 × 103 vs 1.24 × 104, P value = .0155) or prediabetic culture conditions (mean, 7.6 × 103 vs 1.22 × 104, P value = .0059) (Figure 1A). To gain more insight into the decline of CD4 T cell numbers, we labeled the CD3+ CD4 T cells with CFSE, a fluorescent marker used to monitor proliferation by dye dilution, prior to coculture and used flow cytometry to determine exactly how many divisions the T cells underwent in each coculture condition. Our data revealed there was not a difference in the percentage of CD4 T cells that underwent 1–2 cellular divisions (CFSEHIGH) or 3–4 cellular divisions (CFSEINT) between the groups; however, there was a decrease in the percentage of CD4 T cells that completed >5 cellular divisions (CFSELOW) when comparing T cells primed with DCs matured under physiological vs hyperglycemic culture conditions (P value = 0.0229; Figure 1B). This indicates hyperglycemia affects the proliferative capacity of CD4 T cells, resulting in decreased total cell numbers recovered after culture. Another contributing factor for decreased CD4 T-cell numbers when primed with DCs exposed to hyperglycemic culture conditions could be an increase in CD4 T-cell apoptosis. To investigate this, we analyzed caspase-3 expression, a key effector caspase involved in cellular apoptotic pathways,11,12 on CD4 T cells cocultured with DCs primed with different glucose concentrations. Unexpectedly, we found CD4 T cells primed with DCs matured under hyperglycemic culture conditions trended toward decreased caspase-3 expression, indicating decreased apoptotic activity (Figure 1C). Taken together these data suggest that hyperglycemia-matured DCs alter the cellular proliferation and potential survival capacity of the subsequent T cells they prime.
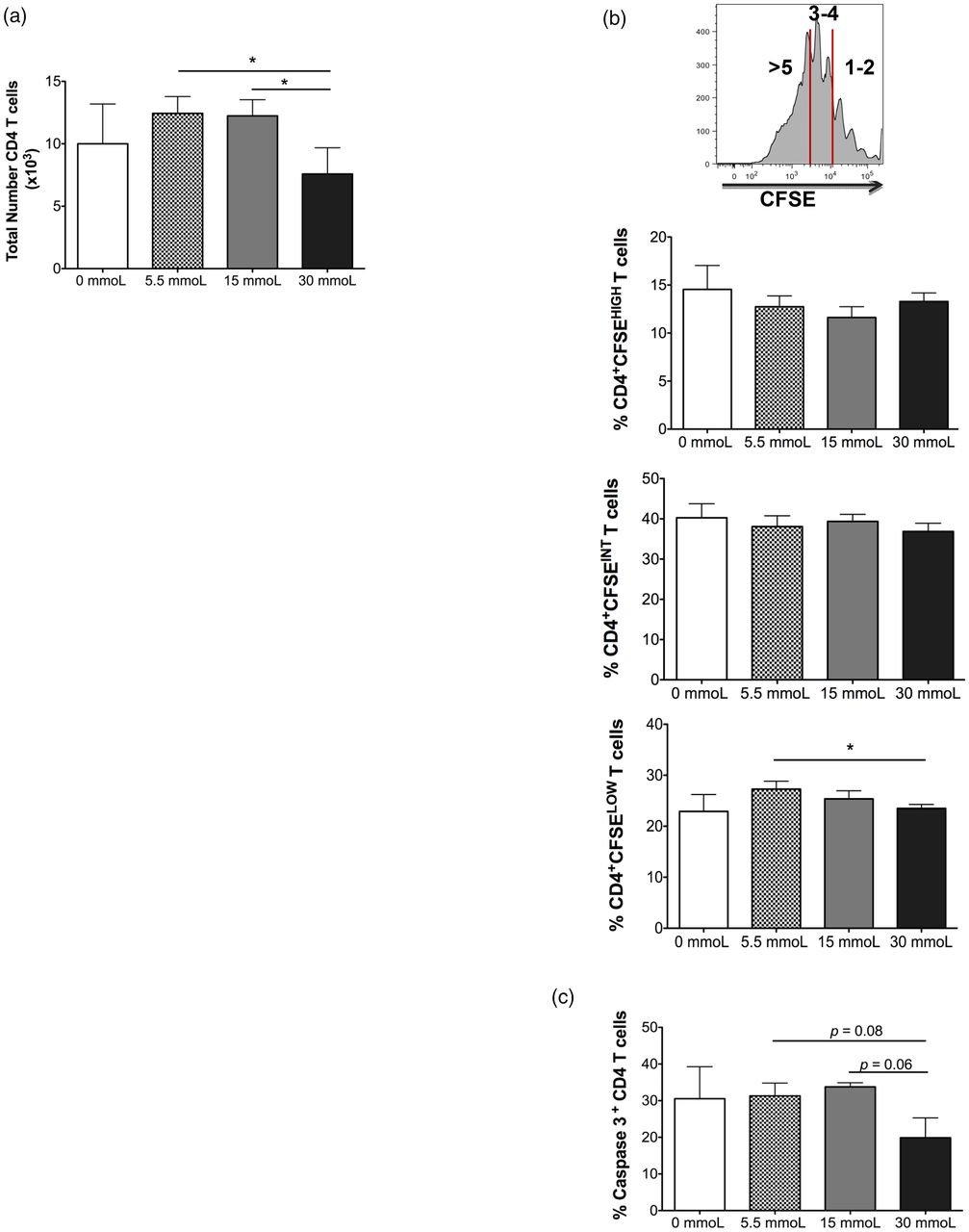
CD4 T cell proliferation after coculture with dendritic cells primed in increasing concentrations of glucose. Peripheral blood–derived CFSE-labeled CD3+ CD4 T cells were cocultured with autologous dendritic cells primed in varying concentrations of glucose. (A) After 7 days in culture, the total CD3+ CD4+ T cells (CD4 T cells) per well were enumerated via flow cytometry. (B) Percentage of CFSE high (top), intermediate (middle), and low (bottom) CD4 T cells was assessed using flow cytometry. CFSE high was defined as 1–2 cellular divisions, CFSE intermediate was defined as 3–4 divisions and CFSE low was defined as >5 cell divisions. (C) Percentage of CD4 T cells expressing caspase-3 was evaluated via flow cytometry. Data represent two independent experiments with 3–6 wells per culture condition. *P = ≦.05
CD4 T Cells Show an Increased Capacity for Mobilization After Coculture With Glucose-stimulated DCs
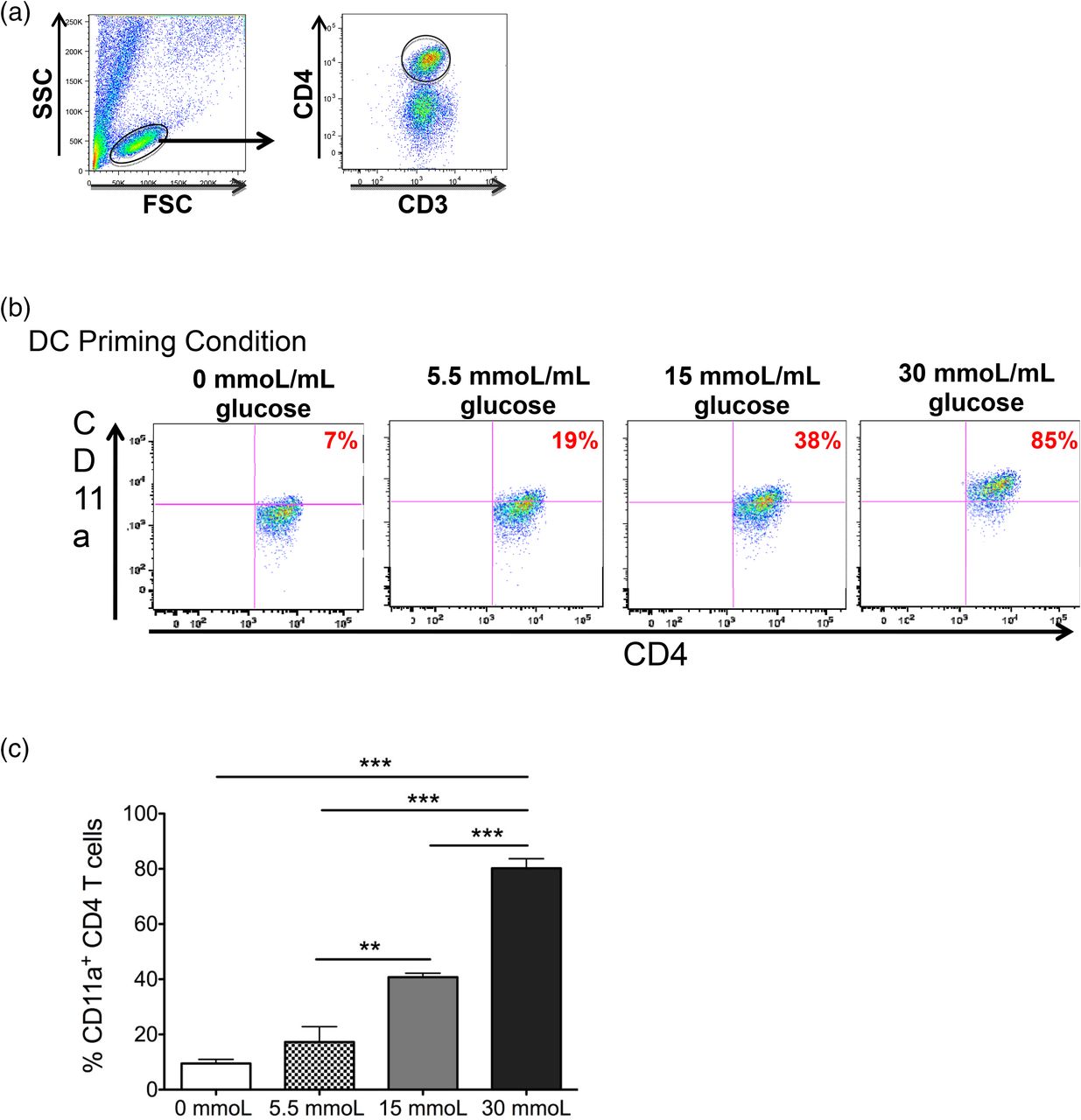
In viral infection models the adhesion molecule CD11a is upregulated on the surface of activated CD4 T cells, allowing them to traffic to sites of inflammation.13,14 In relation to chronic hyperglycemia, it was shown that CD11a plays a crucial role in adipose tissue inflammation through T-cell infiltration and activation.15 However, whether CD11a is upregulated on peripheral blood CD4 T cells in a hyperglycemic environment has not been investigated. Therefore, we examined CD11a expression on the CD4 T cells after 7 days of coculture with DCs primed in different concentrations of glucose. Flow cytometric analysis revealed the expression of CD11a on CD4 T cells cultured with 15 mmol/L (prediabetic) and 30 mmol/L (diabetic) glucose-treated DCs increased significantly compared with CD4 T cells cultured with 5.5 mmol/L (physiological) glucose-treated DCs (Figure 2A–C). Furthermore, this effect was strikingly dose dependent. These results indicate hyperglycemia also affects the phenotype of CD4 T cells, resulting in cells that are more capable of trafficking to areas of inflammation within the body. Taken together, these results indicate CD4 T cells primed by DCs in a hyperglycemic environment display an activated phenotype and support the theory that immune cells, particularly CD4 T cells, play a crucial role in the chronic inflammation observed in patients with type 2 diabetes or prediabetes.
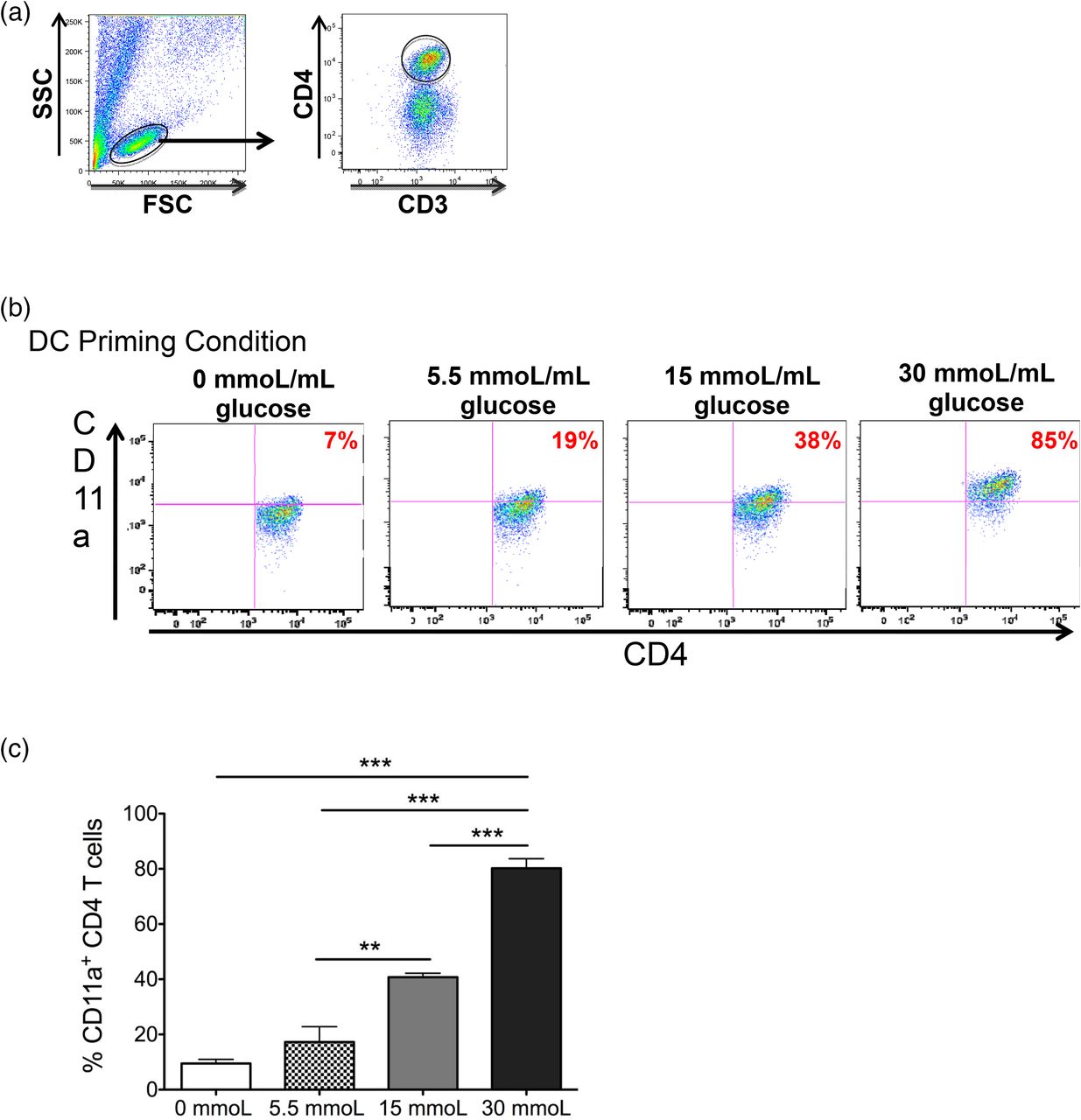
Human CD4 T cells increase their CD11a expression when cultured with DCs primed in increasing concentrations of glucose. Peripheral blood–derived human CD4 T cells were cocultured with autologous dendritic cells primed in varying concentrations of glucose. (A) After 7 days in culture, effector status of CD3+ CD4+ T cells was determined by analysis of CD11a expression using flow cytometry. (B) Representative dot plots of this expression and (C) bar graphs are shown. Data represent two independent experiments with 3–6 wells per culture condition. **P = ≦.01; ***P = ≦.0001
DISCUSSION
Many lines of evidence suggest chronic inflammation not only precedes the onset of prediabetes and type 2 diabetes but is one of the key players in the pathogenesis and progression from obesity and insulin resistance to full onset type 2 diabetes.4,16 Hyperglycemia is the cornerstone of diabetes and has been found to initiate an enhanced inflammatory response by the innate immune system.17 However, the adaptive immune system, specifically T cells, also plays a dominant role in the inflammatory response.18,19 Our data demonstrate that CD4 T cells primed with DCs exposed to the highest amount of glucose (30 mmol/L) have an activated phenotype (increased CD11a) with an altered proliferation profile (decreased CFSE and caspase-3 expression). Through the use of flow cytometry, CD11a was shown to be significantly upregulated on the surface of CD4 T cells cocultured with DCs exposed to hyperglycemic conditions. Although this is in accordance with several clinical studies showing CD4 T cells are activated and play a role in a series of type 2 diabetes–associated complications,20,21 this study provides a specific marker of activation that can be utilized in a clinical setting to monitor patients at risk of developing type 2 diabetes and perhaps stratify these patients in terms of their treatment options.
CD11a is a crucial adhesion molecule that allows cell motility within the body and contributes significantly to lymphocyte’s ability to home to sites of inflammation.22,23 Our study demonstrates the surface expression of CD11a on CD4 T cells significantly increases as the concentration of glucose used to mature DC increases, suggesting hyperglycemia subsequently induces CD4 T cells to be more active and mobile, thereby contributing to the systemic inflammatory process occurring in individuals with prediabetes and type 2 diabetes. In support of this conclusion, CD4 T cells have been shown to promote or sustain the inflammatory process and insulin resistance in metabolic organs of animal models, such as adipose tissue, liver, muscle, and the pancreas. Importantly, our data show a dramatic glucose dose-dependent increase in CD11a expression on CD4 T cells in coculture with DCs. The percentage of CD4 T cells positive for CD11a expression increased in a doubling fashion from 7% to 19%, to 38%, and finally to 85% positive when primed with DCs exposed to no glucose, physiological glucose, prediabetic, and hyperglycemic culture conditions. Furthermore, it might be possible to utilize CD11a as a clinical marker to stratify patients with hyperglycemia who have yet to be diagnosed with prediabetes or full type 2 diabetes. The use of CD11a as an immune marker of activation in the clinical setting could enable differentiation between early preclinical and clinical phases of type 2 diabetes as well as disease complications and progression.
The pattern and frequency of immune subsets present in patients with prediabetes or diagnosed type 2 diabetes remain controversial. Initial studies of T lymphocytes focused on subpopulations present in adipose tissue.7,24 Researchers found that increased numbers of CD4+ Th1 and Th2 cells within the adipose tissue of humans were associated with obesity, inflammation, and insulin resistance. Therefore, we hypothesized that glucose-stimulated DCs would promote CD4 T-cell expansion and survival. Surprisingly, we found hyperglycemic (diabetic) conditions negatively affected the proliferative capacity of CD4 T cells, evident not only by a decrease in total number of cells recovered after coculture but also by the total number of cell divisions CD4 T cells underwent when primed with DCs exposed to hyperglycemic culture conditions. To our knowledge, this is the first study showing the effect of increased glucose on the specific proliferation pattern of human CD4 T cells.
Interestingly, when we analyzed expression of the apoptotic marker caspase-3, we found a trend toward decreased expression on CD4 T cells primed with DCs exposed to diabetic culture conditions, suggesting that apoptosis actually decreases in these cells. In our experimental model, cells were cocultured for 7 days, a relatively short period of time. It is possible if we allowed the coculture to continue past 7 days, we may see an increase in total cell numbers from hyperglycemic culture conditions. This would be in accordance with a study of CD4 T cells in adipose tissue showing an increase in activated CD4 T cells that display characteristics of cellular senescence, defined as a decreased capacity for proliferation while maintaining the ability to produce cytokines.9 Alternatively, it is possible that CD4 T cells primed with DCs matured under hyperglycemic conditions do indeed have a decreased capacity for apoptosis and, additionally, an increased capacity for necrosis. Whether increased necrosis accounts for the decreased total cell numbers in our experimental model is currently under investigation within our laboratory.
In conclusion, we have shown that DCs matured in hyperglycemic conditions subsequently alter the phenotype, proliferative capacity, and survival of CD4 T cells. Moreover, we propose that the observed resistance to apoptosis is through the downregulation of CD27 on the surface of CD4 T cells primed with DCs exposed to diabetic culture conditions. Insights into the immune aspects of type 2 diabetes pathogenesis are invaluable for the design of novel diagnostic and immunotherapeutic strategies to effectively manage this widespread disease. Toward this, we put forward the use of CD11a expression on a patient’s CD4 T cells as a therapeutic marker that is blocked in order to prevent the pathogenic lymphocyte migration to metabolic tissue. Additionally, CD27 could be analyzed to predict progression of prediabetes to type 2 diabetes or to detect the subclinical inflammatory processes that drive this progression. Taken together, the results of this study lend novel insight into potential therapeutic targets for patients with hyperglycemia and diabetes.
FINANCIAL SUPPORT
This work was supported by an American Society of Clinical Laboratory Science (ASCLS) Education and Research Fund (E&R) grant (awarded to N.M.L.) and an SEED grant from Rush University Medical Center, College of Health Sciences (awarded to N.M.L.).
- Received April 2, 2018.
- Accepted May 15, 2018.
American Society for Clinical Laboratory Science
References



{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.